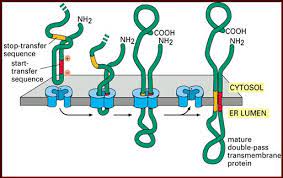
Jalur MAPK mengatur berbagai proses dalam sel, termasuk proses seluler seperti proliferasi, kelangsungan hidup / apoptosis, diferensiasi, perkembangan, kepatuhan, motilitas, metabolisme, dan regulasi gen. Jalur sinyal protein kinase (MAPK) terdiri dari tiga jenis yaitu MAPKKK, MAPKK, dan MAPK. Salah satu modul pensinyalan MAPK yang diidentifikasi dalam sel mamalia adalah melalui jalur GPCR, RTK, dan kedua jalur. Komunikasi sel antara sistem pensinyalan yang berbeda berperan untuk mengkoordinasikan rangsangan ekstraseluler baik dalam kondisi fisiologis maupun patologis. Reseptor permukaan sel merupakan komponen penting dalam proses transmisi sinyal. Protein transmembran terbesar pada permukaan sel adalah reseptor tirosin kinase (RTK) dan reseptor berpasangan G-protein (GPCR)
Aktivasi RTK dicapai dengan mengikat ligan ke domain ekstraseluler, yang dapat menginduksi dimerisasi reseptor dan autofosforilasi. GPCR membentuk ikatan dengan protein G heterotrimerik, yang terdiri dari subunit Gα, Gβ dan Gits. Pengikatan ligan menstabilkan GPCR yang ditempati dalam konformasi pensinyalan aktif di mana protein G heterotrimerik berdisosiasi dalam subunit Gα dan Gβ G yang terikat ke GTP. RTK dan GPCR berperan dalam mengatur proses fisiologis sel. Aktivasi GPCR dapat merangsang aktivitas pensinyalan RTK. Selain itu, RTK menggunakan protein G untuk menginduksi aktivasi jalur dalam sel. Artikel ini bertujuan untuk meninjau protein transmembran untuk komunikasi sel di daerah permukaan, terutama reseptor tirosin kinase (RTK) dan reseptor berpasangan G-protein (GPCR).
Jalur reseptor berpasangan protein G memiliki interaksi dengan PKA-ERK. GCPR akan mengaktifkan reseptor ganda G-protein, protein Gαβ trimerik ke subunit Gα dan mengaktifkan pensinyalan cAMP ke PKA. Gα yang berikatan dengan protein GTP akan mengaktifkan adenylyl cyclase (AC) yang menghasilkan cAMP dari ATP. Selanjutnya, cAMP mengikat subunit PKA dan menginduksi disosiasi holoenzim tersebut. Subunit katalitik (C) kemudian dapat memfosforilasi substrat. Setelah proses selesai, phosphodiesterase (PDE) menghidrolisis cAMP menjadi AMP sehingga jalur cAMP / PKA diaktifkan kembali.
PKA yang sudah aktif dapat merangsang jalur MEK / ERK melalui Rap1 dan B-RAF. Mekanisme ini dimulai dari PKA yang memfosforilasi reseptor β2-adrenergik untuk menghasilkan reseptor β2-adrenergik aktif dari Gα ke Gαi dengan dua konsekuensi yaitu penurunan amplitudo sinyal yang dihasilkan karena Gαi menghambat adenilat siklase, dan pengaturan jalur ERK1 / ERK2 melalui Gβγ, SRC RAS, dan RAF-1. cAMP menstimulasi sinyal ke protein Epac dan Rit ke MEK / ERK. Konsentrasi cAMP intraseluler diatur oleh dua kelas enzim dengan efek berlawanan. Adenylyl cyclase menghasilkan cAMP dari ATP, sedangkan cAMP phosphodiesterases (PDE) menghidrolisis cAMP menjadi AMP.
Mekanisme dimana sel dapat membentuk ikatan antara ERK2 dan PKA melalui interaksi langsung antara ERK2 dan PDE4. Interaksi antara ERK2 dan PDE4 membutuhkan kekhususan lokasi untuk FQF dan KIM docking. Jadi aktivasi ERK2 menghambat atau merangsang pensinyalan cAMP / PKA tergantung pada subtipe PDE4. Kesimpulannya, jalur GCPR ini dapat menggunakan aktivasi Epac1 yang diinduksi cAMP dan selanjutnya Rap1, B-RAF, Rit dan MEK1 / 2 untuk mengaktifkan ERK1 / 2, atau aktivasi cAMP / Epac1 yang menghasilkan protein SSR5.
Dalam banyak kasus, RTK dan GPCR membentuk kompleks di mana sinyal dapat diintegrasikan untuk menghasilkan stimulasi dan jalur intraseluler yang lebih efisien. PDGF dan sphingosine 1-phosphate (S1P) menginduksi RTK ke kompleks reseptor 1-fosfat PDGFRβ-sphingosine 1 (S1P1). Ini mendorong fosforilasi protein pengikat tirosin terkait Grb2 1 (Gab1), yang bergantung pada aktivasi protein G dan RTK. Selanjutnya akan mengaktifkan cAMP, PKA untuk menghasilkan protein RAS, B-RAF, dan c-RAF-1 yang menghasilkan MEK dan ERK (Cattaneo et al, 2014). Kesimpulannya, banyak protein yang berperan baik pada jalur GPCR maupun RTK, yaitu Grb2, AC, cAMP, PKA, RAS, B-RAF, dan c-RAF-1, menghasilkan MEK dan ERK1.
Protein transmembran terbesar di permukaan sel untuk komunikasi di permukaan adalah reseptor tirosin kinase (RTK) dan reseptor berpasangan G-protein (GPCR). RTK dan GPCR berperan dalam mengatur proses fisiologis sel. Aktivasi GPCR dapat merangsang aktivitas pensinyalan RTK. Selain itu, RTK menggunakan protein G untuk menginduksi aktivasi jalur dalam sel.
Penulis: Nanda Rachmad Putra Gofur drg,.M.Biomed
Informasi detail dari riset ini dapat dilihat pada tulisan kami di:
Nanda Rachmad Putra Gofur, Aisyah Rachmadani Putri Gofur, Soesilaningtyas, Rizki Nur Rachman Putra Gofur, Mega Kahdina and Hernalia Martadila Putri (2021) The Transmembrane Proteins for Cell Communication in Surface Region: A Review Article. Archives of Clinical Case Reports 2: 156-158.